Xác thực các tác nhân gây bệnh WFS
Sự đồng nhiễm với V. fluvialis (Vf), V. coralliilyticus (Vc) và V. tubiashii (Vt) đã gây ra các triệu chứng điển hình của WFS, chẳng hạn như chậm chạp, giảm ăn uống, ruột teo và tỷ lệ tử vong cao (Hình 1b, c). Tuy nhiên, việc nhiễm một dòng Vibrio đơn lẻ hay các dòng Vibrio kết hợp giữa Vf, Vc và Vt không thể tái hiện các hội chứng này (Hình bổ sung 1). Thêm vào đó, ba tác nhân Vibrio được phát hiện trong ruột, đáng kể hơn ở tôm nhiễm NV (Vibrio nhiễm mà không bổ sung probiotics) so với tôm đối chứng (CK) (Hình 1d). Tất cả những điều này cho thấy ba tác nhân Vibrio đã được phân lập từ tôm nhiễm WFS, phối hợp gây ra triệu chứng WFS, và gia tăng ở các cá thể tái nhiễm, điều này hoàn toàn thỏa mãn các quy tắc của Koch. Do đó, Vf, Vc và Vt là các tác nhân gây bệnh của WFS. Tuy nhiên, chúng tôi không thể hoàn toàn loại trừ khả năng ảnh hưởng của các tác nhân gây bệnh khác trong WFS ở tôm. Cần có thêm công việc để giải quyết nguyên nhân gây ra WFS.
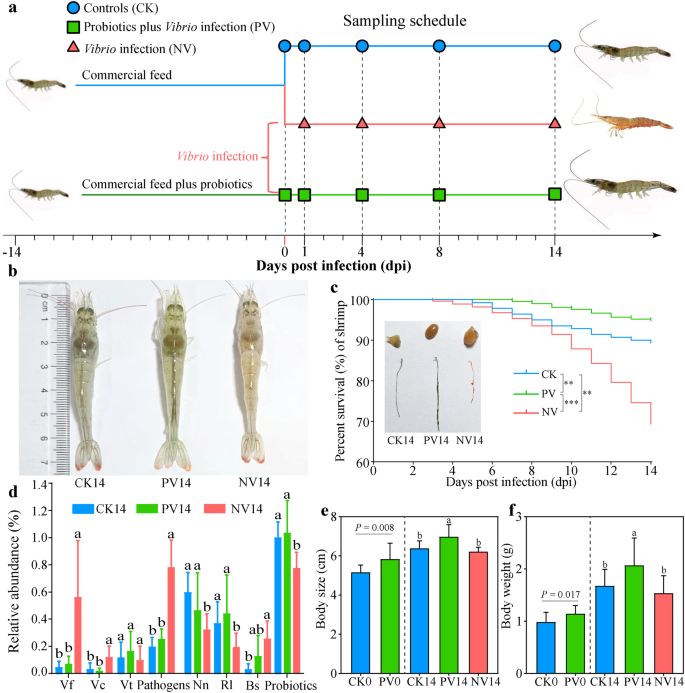
a Thiết kế thí nghiệm. Tôm được cho ăn một chế độ ăn cơ bản có bổ sung/probiotics đối kháng (1 × 107 CFU/g chế độ ăn) trong 14 ngày, và sau đó bị nhiễm bằng cách tắm với Vibrio fluvialis (Vf), V. coralliilyticus (Vc) và V. tubiashii (Vt) (với tỷ lệ 7:2:1) với mật độ 1 × 108 CFU/mL. b Các kiểu hình tôm giữa các điều trị 14 ngày sau nhiễm (dpi). c Đồ thị Kaplan-Meier về tỷ lệ tử vong của tôm theo dpi. ** và *** cho thấy sự khác biệt có ý nghĩa giữa các điều trị khác nhau ở P P d So sánh sự phong phú tương đối (mean ± độ lệch chuẩn) của ba tác nhân, tổng số tác nhân, các dòng đối kháng được phát hiện, Ruegeria lacuscaerulensis (Rl), Nioella nitratireducens (Nn), Bacillus subtilis (Bs), và tổng số probiotics trong ruột giữa các điều trị vào 14 dpi. e Chiều dài cơ thể tôm (mean ± độ lệch chuẩn), f Trọng lượng tôm (mean ± độ lệch chuẩn) vào 0 dpi (trước khi nhiễm) sử dụng kiểm định t không đối cặp, và vào 14 dpi sử dụng phân tích phương sai một chiều (ANOVA). Các chữ cái thường khác nhau chỉ ra sự khác biệt có ý nghĩa giữa các điều trị.
Thiết kế probiotics đối kháng chống lại WFS
Xem xét rằng các loài chiếc khóa là mục tiêu tuyệt vời cho các can thiệp dựa trên vi khuẩn đường ruột14,15, chúng tôi đã xác định sự kết hợp tối ưu của các loài chiếc khóa với điểm can thiệp kết hợp (CIS) lớn nhất bằng cách sử dụng Phương pháp loại trừ đặc trưng lặp lại (IFE). IFE, một chiến lược lựa chọn đặc trưng dựa trên thuật toán tham lam22, được sử dụng để tìm kiếm sự kết hợp tối ưu của các loài vi sinh vật cho can thiệp. CIS đại diện cho hiệu ứng can thiệp sau khi can thiệp đồng thời của sự kết hợp tối ưu của các loài vi sinh vật, do đó giá trị CIS cao hơn cho thấy hiệu ứng can thiệp tốt hơn15. Bốn loài chiếc khóa hàng đầu từ vi khuẩn đường ruột của tôm WFS đã đóng góp một CIS là 0.807 (Hình 2a và Bảng bổ sung 1). Nhắm mục tiêu vào bốn loài chiếc khóa trong tôm WFS đã kích thích sự phục hồi ở các thành viên là loài trung tâm trong vi khuẩn đường ruột của tôm khỏe mạnh (Hình 2b). Do đó, bốn loài chiếc khóa này đã phục hồi tối đa về một vi sinh vật đường ruột khỏe mạnh và được chọn làm probiotics đối kháng để ngăn ngừa WFS.
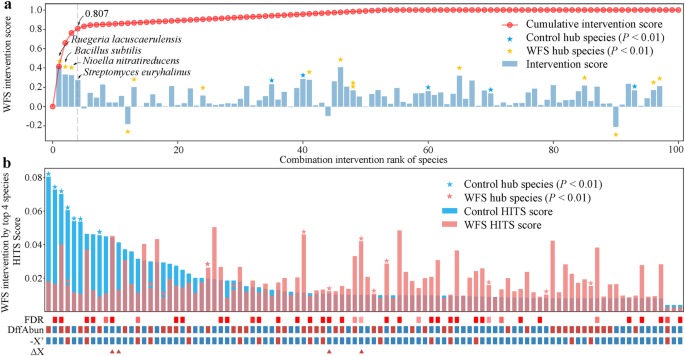
a Các điểm can thiệp (IS) của 100 cộng sinh đường ruột hàng đầu. IS của mỗi loài được trình bày trong biểu đồ cột và các ngôi sao với màu xanh lam và đỏ cho thấy độ quan trọng của điểm số Tìm kiếm chủ đề do siêu liên kết tác động (HITS) trong vi khuẩn đường ruột của tôm khỏe mạnh và tôm WFS, tương ứng. IS âm có nghĩa là sự thay đổi khỏi vi khuẩn đường ruột của tôm khỏe mạnh. Đường cong đỏ biểu thị các điểm can thiệp kết hợp (CIS) của các vi sinh vật được chọn lần lượt bởi DIS, điều này đại diện cho hiệu ứng can thiệp sau khi can thiệp đồng thời của nhiều loài. Bốn loài chiếc khóa hàng đầu, đạt được CIS > 0.8, được chỉ định. b Tác động của sự can thiệp vi sinh vật đến vi khuẩn đường ruột WFS theo DIS với bốn loài đối kháng. Các điểm số HITS của các loài trong vi khuẩn đường ruột của tôm khỏe mạnh được sắp xếp trong biểu đồ cột, và các loài trung tâm được đánh dấu bằng các ngôi sao. DiffAbun: sự thay đổi độ phong phú từ tôm khỏe mạnh sang tôm WFS (đỏ: tăng, xanh lam: giảm), với tỷ lệ phát hiện giả sai (FDR) được chỉ ra ở trên (đỏ: FDR X′: đại diện âm cho sự thay đổi độ phong phú vi khuẩn tức thì ngay sau can thiệp (đỏ: −X′ > 0, xanh lam: −X′ X.
Tỷ lệ tử vong ở tôm NV xảy ra vào 3 ngày sau nhiễm (dpi) và sau đó. Ngược lại, nhờ việc bổ sung probiotics trong 14 ngày, tỷ lệ tử vong đầu tiên ở tôm PV (probiotics cộng với nhiễm Vibrio) được ghi nhận vào 7 dpi, trì hoãn 4 ngày so với tôm NV (Hình 1a, c). Đặc biệt, tỷ lệ tử vong tích lũy ở tôm PV là 5.2% vào 14 dpi, thấp hơn đáng kể so với tôm CK (10.7%) hoặc NV (30.4%) (Hình 1c). Các probiotics đối kháng được phân lập từ tôm khỏe mạnh, do đó đã được phát hiện trong nhóm CK với độ phong phú tương đối cao (Hình 1d). Như mong đợi, các mối tương quan tiêu cực và có ý nghĩa đã được phát hiện giữa tổng độ phong phú tương đối của các probiotics đối kháng và của ba tác nhân Vibrio (Hình bổ sung 2). Ngoài ra, việc bổ sung probiotics đã làm tăng hoạt động pepsin và lipase của tôm vào 0 dpi (Hình bổ sung 3a, b), và do đó cải thiện trọng lượng và chiều dài cơ thể so với tôm CK (Hình 1e, f). Sau 14 ngày nhiễm, tôm PV có hoạt động miễn dịch và tiêu hóa cao hơn đáng kể so với tôm NV, ngoại trừ phosphatase acid (Hình bổ sung 3). Tóm lại, các probiotics mà chúng tôi thiết kế bảo vệ hiệu quả tôm khỏi WFS, cải thiện tỷ lệ sống sót, miễn dịch và sản xuất của tôm.
Phản ứng của cộng đồng vi khuẩn theo thời gian sau nhiễm
Hứng thú với các kiểu hình khác nhau (Hình 1), chúng tôi đã tìm cách làm sáng tỏ bất kỳ sự khác biệt nào về thành phần trong vi khuẩn đường ruột giữa các điều trị. Sau khi làm hiếm hóa đến 16,019 chuỗi mỗi mẫu, có 2195 biến thể chuỗi amplicon (ASVs) trên 120 mẫu đã được đăng ký (Bảng bổ sung 2). Cả việc bổ sung probiotics và nhiễm tác nhân đều đã làm thay đổi đáng kể vi khuẩn đường ruột tôm và cộng đồng vi sinh vật (Hình bổ sung 4a, b). Tuy nhiên, việc bổ sung probiotics đã phần nào chống lại tác động của nhiễm tác nhân đến vi khuẩn đường ruột, điều này được hỗ trợ bởi sự tách biệt dần dần dọc trục 1 (Hình bổ sung 4a). Tương ứng, sự khác biệt nổi bật trong độ phong phú của các chi thống trị đã được quan sát giữa tôm NV và CK, với sự thay đổi vừa phải từ các chi này ở tôm PV vào 14 dpi (Hình bổ sung 5). Ba trong bốn dòng đối kháng được phát hiện trong ruột, với độ phong phú tương đối ở tôm NV thấp hơn đáng kể so với ở tôm CK hoặc PV vào 14 dpi (Hình 1d). Mặc dù Streptomyces euryhalinus không thể phát hiện vào 14 dpi (Hình 1d), độ phong phú của chi Streptomyces trong cộng đồng vi khuẩn ruột của tôm PV cao hơn đáng kể so với NV (Hình bổ sung 5a). Tương ứng, hiệu quả của probiotics kéo dài hơn cả khả năng sống sót của chúng, thay vào đó, vật chủ có thể hưởng lợi từ các chuyển hóa và thành phần tế bào của probiotics23. Phân tích phương sai đa biến theo từng phép (PERMANOVA) đã chỉ ra rằng dpi, probiotics và nhiễm bệnh đã giải thích 6.3%, 4.7% và 5.2% (P 3).
Tốc độ thay đổi theo thời gian của vi khuẩn đường ruột ở tôm CK (độ dốc = −0.028) thấp hơn đáng kể (P = 0.044, phân tích phương sai hai chiều) so với tôm NV (độ dốc = −0.060) (Hình bổ sung 4c), cho thấy việc nhiễm tác nhân đã thúc đẩy việc thay thế các loài vi khuẩn đường ruột theo sự tiến triển của WFS. Tuy nhiên, tốc độ thay đổi theo thời gian đã bị giảm đi bởi việc bổ sung probiotics, với độ dốc thấp nhất ở tôm PV (độ dốc = −0.007) (Hình bổ sung 4c). Sự ổn định của vi khuẩn đường ruột được đánh giá bằng độ biến đổi trung bình (AVD). Giá trị AVD thấp hơn cho thấy ổn định cộng đồng cao hơn24. Giá trị AVD tương đương giữa tôm PV và CK vào 0 dpi (Hình bổ sung 4d), cho thấy rằng sự định cư của probiotics không làm gián đoạn sự ổn định của vi khuẩn đường ruột của tôm. Ngược lại, AVD ở tôm PV thấp hơn đáng kể so với nhóm NV vào 14 dpi (Hình bổ sung 4d), do đó các probiotics đối kháng đã chống lại sự không ổn định của vi khuẩn đường ruột do nhiễm bệnh gây ra. Tương tự, suy luận từ Bugbase cho thấy việc nhiễm tác nhân đã làm tăng tiềm năng của các vi sinh vật kị khí, hình thành biofilm và các tác nhân nghi ngờ trong tôm NV so với tôm CK (Hình 3e−g). Đặc biệt, tiềm năng của các tác nhân nghi ngờ là thấp nhất ở tôm PV trong ba nhóm (Hình 3g).
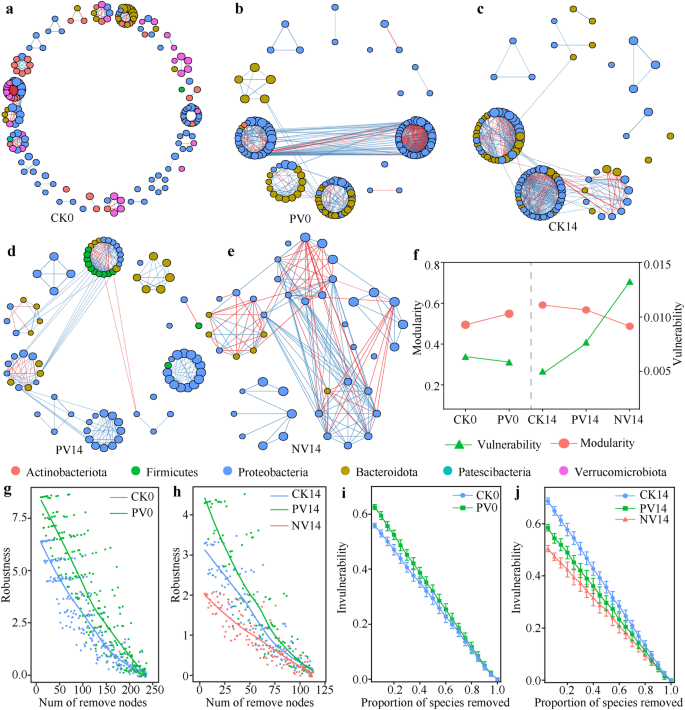
Mạng lưới đồng xuất hiện cho a CK (tôm đối chứng), b PV (probiotics cộng với nhiễm Vibrio) tôm vào 0 dpi, c CK, d PV, e NV (Vibrio nhiễm mà không bổ sung probiotics) tôm vào 14 dpi, tương ứng. Mỗi nút (điểm) đại diện cho ASV cá nhân, được tô màu theo phylum liên quan. Kích thước của một nút đại diện cho độ phong phú tương đối của nó trong mỗi cộng đồng vi khuẩn. Các cạnh đại diện cho các mối tương quan Spearman có ý nghĩa (|r| > 0.8 và P f cấu trúc và độ dễ bị tổn thương, g độ ổn định vào 0 dpi, h độ ổn định vào 14 dpi, i độ không thể tổn thương vào 0 dpi, j độ không thể tổn thương vào 14 dpi, tương ứng.
Các quá trình sinh thái điều khiển vi khuẩn đường ruột tôm
Sự tăng cao của tính ngẫu nhiên điều khiển vi khuẩn đường ruột của vật chủ tạo điều kiện cho sự xâm nhập của các tác nhân bên ngoài25, vì vậy chúng tôi đã nâng cao xem việc bổ sung probiotics có tăng cường tầm quan trọng của nguyên tắc điều chỉnh phía dưới vi khuẩn đường ruột tôm không, bằng cách sử dụng mô hình cộng đồng trung tính (NCM)26. NCM giả định rằng cộng đồng vi sinh vật là nguồn loài cho vi khuẩn đường ruột tôm. Sự phù hợp tổng thể với NCM ở tôm CK (R2 = 0.777, Hình bổ sung 6a) thấp hơn so với tôm NV (R2 = 0.786, Hình bổ sung 6c), do đó việc nhiễm tác nhân đã làm tăng tính ngẫu nhiên điều khiển vi khuẩn đường ruột tôm. Tuy nhiên, việc bổ sung probiotics đã làm giảm tác động của nhiễm bệnh, với giá trị R2 là 0.726 ở tôm PV (Hình bổ sung 6b). Một cách đồng nhất, 93.5% tổng số ASVs được phân bố một cách trung tính giữa vi khuẩn đường ruột tôm NV và nước nuôi của chúng, tỷ lệ này cao hơn nhiều so với tỉ lệ trong tôm PV (50.5%) và CK (50.1%). Ngược lại, tỷ lệ ASVs quá đại diện hoặc quá thiếu đại diện có xu hướng ngược lại (Hình bổ sung 6a−c).
Chúng tôi đã đi sâu vào các quá trình sinh thái thành năm quá trình sinh thái bằng cách sử dụng mô hình iCAMP (Hình bổ sung 6d−f). Tầm quan trọng tương đối của lựa chọn đồng đều điều khiển vi sinh vật đường ruột ở tôm CK là 33.9% (Hình bổ sung 6d), điều này giảm xuống 24.4% ở tôm NV (Hình bổ sung 6f). Ngược lại, sự đóng góp của lựa chọn đồng đều đã tăng lên mạnh mẽ đến 60.2% ở tôm PV (Hình bổ sung 6e). Ngược lại, quá trình trôi nổi đã nổi lên như một quá trình sinh thái chủ yếu ở tôm NV (62.2%), trong khi quá trình này giảm xuống 17.8% ở tôm PV (Hình bổ sung 6e, f).
Tác động của probiotics và nhiễm bệnh đến mạng lưới của vi khuẩn đường ruột
Để xác định tác động của probiotics và nhiễm bệnh đến các tương tác sinh học giữa các vi sinh vật đường ruột, chúng tôi đã xây dựng các mạng lưới đồng xuất hiện cho từng nhóm vào 0 dpi và 14 dpi, tương ứng (Hình 4). Tôm PV thể hiện độ ổn định mạng cao hơn so với tôm CK vào 0 dpi, với sự hỗ trợ của độ dễ bị tổn thương thấp hơn, độ modularity cao hơn và độ bền cao hơn (Hình 4). Tuy nhiên, nhiễm tác nhân đã làm giảm đáng kể độ bền của mạng lưới so với tôm CK vào 14 dpi. Tác động làm mất ổn định này đã được đảo ngược bởi việc bổ sung probiotics, với độ bền cao nhất ở tôm PV (Hình 4h). Sau khi loại bỏ ngẫu nhiên một số nút, tỷ lệ các loài còn lại trong mạng lưới của tôm PV liên tục cao hơn tỷ lệ của tôm CK (Hình 4i), cho thấy độ không thể tổn thương của mạng lưới đã được củng cố bởi việc bổ sung probiotics. Một lần nữa, các probiotics đối kháng đã trung hòa độ không thể tổn thương của mạng lưới, modularity và độ bền bị phá vỡ bởi nhiễm tác nhân (Hình 4f, j).
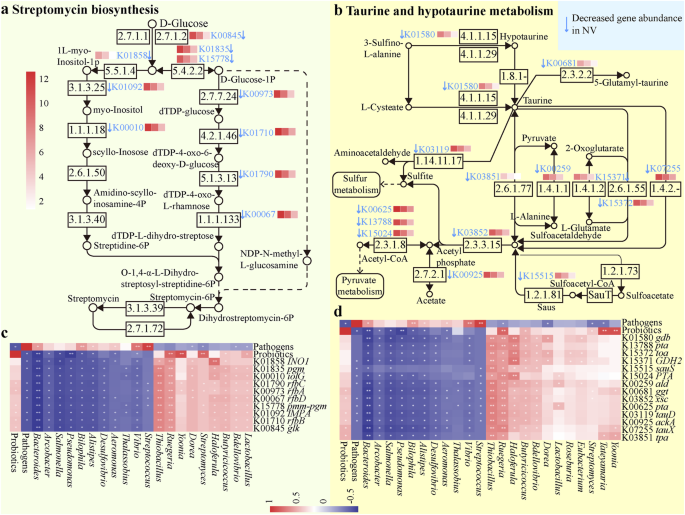
a Tổng hợp streptomycin. b Chuyển hóa taurine và hypotaurine. Các mũi tên hướng xuống cho thấy các con đường chức năng khác nhau bị suy giảm ở tôm NV. Các chi vi khuẩn đường ruột, tổng độ phong phú của các tác nhân và tổng độ phong phú của probiotics thể hiện các mối liên quan mạnh mẽ với các gen liên quan đến c tổng hợp streptomycin, d chuyển hóa taurine và hypotaurine. Chỉ các mối tương quan hạng Spearman với P
Tác động của probiotics lên tiềm năng chức năng chống nhiễm bệnh
Để hiểu rõ hơn về tác động tích cực của probiotics đối với tương tác giữa tôm và vi khuẩn đường ruột chống lại WFS, chúng tôi đã khám phá sự khác biệt về tiềm năng chức năng giữa ba điều trị. Sau khi kiểm soát chất lượng, 171.2 GB các chuỗi chất lượng cao đã được tạo ra cho 18 vi khuẩn đường ruột tôm vào 14 dpi, trong số đó 92.7% các chuỗi metagenomic vượt qua ngưỡng Q30 (Bảng bổ sung 4). So với tôm CK, độ phong phú của 163 con đường, bao gồm chuyển hóa Butanoate map00650, chuyển hóa Propanoate map00640 và tổng hợp axit béo map00061, đã giảm đáng kể (P t test) ở tôm NV vào 14 dpi. Ngược lại, 57 đường dẫn, chẳng hạn như thụ thể Toll-like (TLR) map04620, mTOR map04150 và các con đường tín hiệu JAK-STAT map04630, thể hiện xu hướng ngược lại (Hình bổ sung 7a). Tương tự, 39 đường dẫn cung cấp tổng hợp streptomycin map00521, tổng hợp axit mật thứ cấp map00121 và chuyển hóa Butanoate map00650 giảm đáng kể (P t test), trong khi 6 đường dẫn như chuyển hóa Propanoate map00640 và thụ thể lectin loại C (CLR) tín hiệu map04625, đã tăng cường ở tôm NV so với tôm PV vào 14 dpi (Hình bổ sung 7b). Tuy nhiên, chỉ có ba đường dẫn khác biệt được phát hiện giữa tôm CK và PV (Hình bổ sung 7c), do đó, cấu trúc chức năng của vi khuẩn đường ruột giữa hai nhóm là tương đương. Nói cách khác, việc bổ sung probiotics đã giảm thiểu đáng kể các tác động tiêu cực do nhiễm bệnh gây ra.
Có những mối liên hệ chặt chẽ giữa các tác nhân, probiotics và vi khuẩn đường ruột (Hình bổ sung 7d, e). Các tác nhân Vibrio kích thích các con đường thúc đẩy phản ứng viêm, chẳng hạn như NF-κB map04064, TLR map04620, thụ thể giống NOD (NLR) map04621 và các con đường tín hiệu CLR map04625. Ngược lại, probiotics có tác động tích cực đến các con đường trao đổi chất chủ chốt đa dạng, bao gồm taurine và hypotaurine map00430, pyruvate map00620, propanoate map00640 và butanoate map00650 (Hình bổ sung 7d, e). Đặc biệt, ba tác nhân Vibrio tương tác tích cực với nhau (Hình bổ sung 7e), củng cố vai trò tương tác của chúng trong nguyên nhân của WFS ở tôm.
Ấn tượng, bằng cách căn chỉnh các gen bị thay đổi với cơ sở dữ liệu KEGG, chúng tôi đã cấu trúc các con đường liên quan đến tổng hợp streptomycin (Hình 3a), chuyển hóa taurine và hypotaurine (Hình 3b), chuyển hóa propanoate (Hình bổ sung 8), và chuyển hóa butanoate (Hình bổ sung 9). Độ phong phú của các gen có liên quan này giảm mạnh ở tôm NV so với các cá thể CK. Tuy nhiên, mức độ và mức độ nghiêm trọng của các giảm sút này ít rõ rệt hơn ở tôm PV so với tôm NV. Đáng chú ý, độ phong phú của các gen tham gia có mối liên hệ tích cực với các chi có lợi như Bdellovibrio, Butyricicoccus, Eubacterium, Clostridium và Streptomycete, và tổng độ phong phú của probiotics, trong khi bị tác động tiêu cực bởi các chi gây hại Vibrio, Pseudoalteromonas và Streptococcus, và tổng độ phong phú của ba tác nhân Vibrio (Hình 3, Hình bổ sung 8 và 9).
Các transcriptome của tôm bị thay đổi đáng kể thiết yếu cho khả năng kháng WFS
Xem xét cuộc tranh luận rằng độ phong phú của mRNA không tương đương với hoạt động, do đó hoạt động của lipase, lysozyme hoặc phosphatase kiềm đã được hồi quy với mức độ mRNA của gen mã hóa tương ứng, tương ứng. Như mong đợi, một mối liên hệ tích cực và có ý nghĩa đã được phát hiện giữa hoạt động enzyme và biểu hiện của gen mã hóa phù hợp (Hình bổ sung 10). Do đó, các hồ sơ biểu hiện bị thay đổi ở tôm có thể, ít nhất phần nào, chỉ ra các hoạt động. Phân tích transcriptome tiết lộ rằng 55 và 7 gen đã được biểu hiện cao hơn và thấp hơn ở tôm PV so với tôm CK vào 0 dpi, tương ứng (Hình 5a). Việc bổ sung probiotics đã kích thích đáng kể các gen liên quan đến tổng hợp các chuyển hóa phụ, pyruvate và glutathione, thực bào trung gian FcγR, và điều chỉnh cytoskeleton actin (Hình 5e). Nhiễm tác nhân đã làm biến đổi các hồ sơ biểu hiện một cách đáng kể, trong đó 42 và 142 gen đã được biểu hiện cao hơn và thấp hơn ở tôm NV so với tôm CK vào 14 dpi (Hình 5b). Đối với các gen được biểu hiện cao hơn ở tôm NV, chúng tôi đã rõ ràng quan sát được sự phong phú trong các con đường tín hiệu và miễn dịch. Ngược lại, các con đường chuyển hóa đã bị suy giảm ở tôm NV vào 14 dpi, chẳng hạn như các chuyển hóa tryptophan và pyruvate (Hình 5f). Tuy nhiên, các biểu hiện bị kích thích bởi nhiễm bệnh đã bị chống lại bởi việc bổ sung probiotics, như được hỗ trợ bởi 10 gen được biểu hiện cao hơn và 27 gen được biểu hiện thấp hơn ở tôm NV so với tôm PV vào 14 dpi (Hình 5c). Ấn tượng, sự phong phú do probiotics kích thích của chuyển hóa pyruvate, tự thực bào, và glycolysis/gluconeogenesis vẫn có thể phát hiện ở tôm PV vào 14 dpi (Hình 5g). Tuy nhiên, chúng tôi cũng đã xác định được 13 gen được biểu hiện cao hơn và 16 gen được biểu hiện thấp hơn ở tôm PV so với tôm CK. Cụ thể, các con đường tín hiệu cGMP-PKG và AMPK đã được kích thích ở tôm PV, trong khi chuyển hóa cysteine và methionine, con đường tín hiệu chemokine và hệ thống bài tiết vi khuẩn cho thấy xu hướng ngược lại vào 14 dpi (Hình 5d, h). Một cách đồng nhất, việc bổ sung probiotics đã cải thiện sự biểu hiện của các gen miễn dịch (Hình bổ sung 11a, b), và giảm thiểu phản ứng viêm (11c, d). Đặc biệt, độ phong phú của các gen được biểu hiện khác nhau trong đường ruột (DEGs) có mối liên hệ mạnh mẽ với các con đường chức năng khác nhau (DFPs) ở vi khuẩn đường ruột (Hình bổ sung 12), cho thấy sự giao tiếp giữa microbiota đường ruột và miễn dịch của tôm.
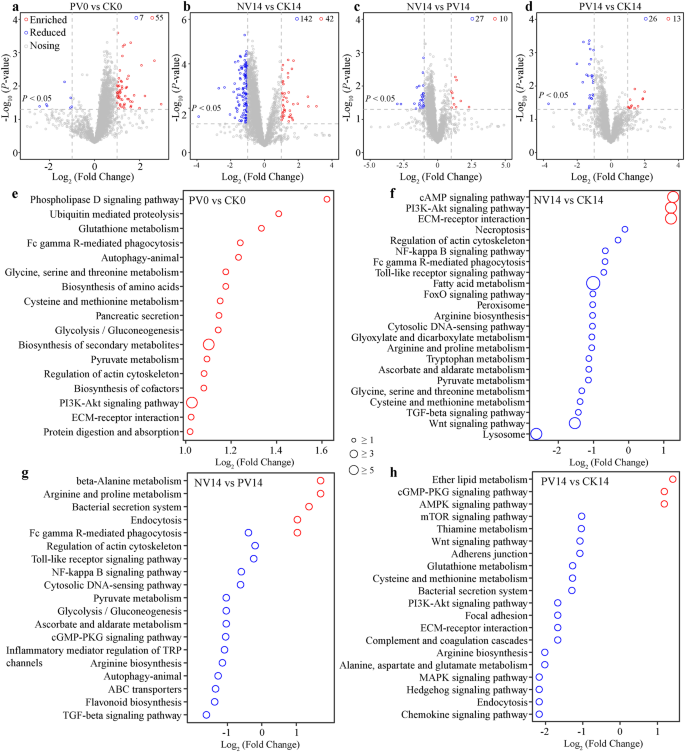
Các đồ thị núi lửa mô tả sự phân bố của các gen biểu hiện khác nhau (DEGs, a–d) và các con đường tương ứng của chúng (e–g) giữa tôm PV so với CK (a hoặc e) vào 0 dpi, NV so với CK (b hoặc f), NV so với PV (c hoặc g), PV so với CK (d hoặc h) vào 14 dpi sử dụng kiểm định t hai đầu không đối cặp, tương ứng. Các số liệu chỉ ra số lượng DEGs được biểu hiện cao hơn và thấp hơn. Các vòng tròn màu đỏ và xanh lam lần lượt chỉ ra các con đường được biểu hiện cao hơn hoặc thấp hơn, trong khi đường kính tỷ lệ thuận với số lượng DEGs đã phát hiện trong một con đường nhất định.
Các probiotics đối kháng cải thiện sự biểu hiện của các gen nối chặt đường ruột
Việc bổ sung probiotics đã kích thích đáng kể (P t test) mức độ biểu hiện của mucin 6, mucin 2, protein kích hoạt GTPase Rho 17 (ARHGAP17), protein liên quan đến Ras Rab-8A (RAB8A) và thành viên của phân họ MAGUK p55 số 5 (MPP5) (Hình bổ sung 13). Tuy nhiên, việc nhiễm tác nhân đã làm giảm đáng kể biểu hiện của mucin 2, ARHGAP17, RAB8A và MPP5. Ngược lại, các probiotics đối kháng của chúng tôi hiệu quả duy trì hoặc thậm chí cải thiện biểu hiện của các gen nối chặt đường ruột này (Hình bổ sung 13).
Phân tích tích hợp giữa các tác nhân, probiotics, microbiome đường ruột và transcriptome của tôm
Chúng tôi đã tích hợp nhiều dữ liệu omics để xác định các đặc điểm phân tử liên quan sinh học và đáng tin cậy giữa microbiome đường ruột tôm và transcriptome. Các đồ thị mẫu của mô hình DIABLO (Phân tích Tích hợp Dữ liệu cho Khám Phá Đặc Trưng bằng cách sử dụng phương pháp thành phần tiềm ẩn cho Omics) cho thấy rằng transcriptome tôm và microbiome đường ruột phân biệt hiệu quả ba nhóm tôm (Hình 6a−c). Các thành phần tiềm ẩn của mỗi bộ dữ liệu omic có mối tương quan cao (r = 0.91 − 0.97) giữa chúng (Hình 6d), tiết lộ sự đồng nhất tốt giữa ba bộ dữ liệu ở cấp mẫu. Mối tương quan giữa các thành phần của mỗi bộ dữ liệu đã được tối đa hóa như đã chỉ định trong ma trận thiết kế. Ba loại dữ liệu, đại diện cho các cấp độ khác nhau, thể hiện một mối tương quan cao ở cấp độ thành phần (Hình 6e). Dựa trên chữ ký phân tử đa omics, 18 mẫu đã được phân cụm theo điều trị mà các mẫu thuộc về (Hình 6f). Bản đồ circosPlot trực quan hóa mối tương quan và tính tương tác của các dữ liệu OMIC khác nhau bằng cách sử dụng 14 chi vi khuẩn, 5 con đường KEGG trong microbiota đường ruột và 15 gen chức năng của tôm, với ngưỡng hệ số tương quan là 0.5, thể hiện sự khác biệt rõ rệt giữa các điều trị ghép (Hình 6g).
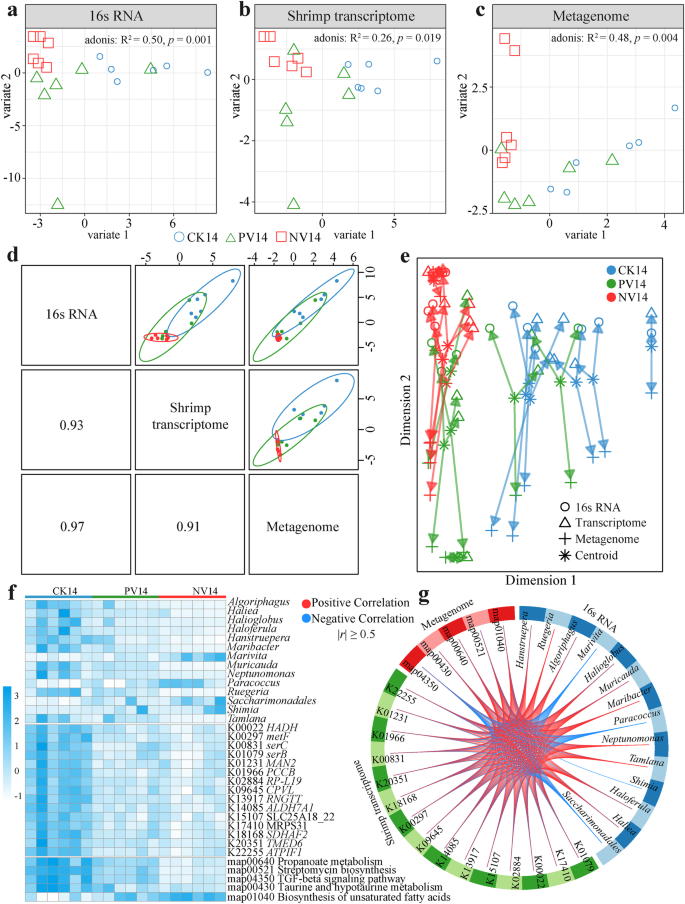
Các đồ thị mẫu cho a thành phần, b cấu trúc chức năng của microbiota đường ruột, và c transcriptome đường ruột của tôm. d Biểu đồ phân tán mẫu hiển thị thành phần đầu tiên trong mỗi bộ dữ liệu (biểu đồ chéo trên) và mối tương quan Pearson giữa từng thành phần (biểu đồ chéo dưới). e Biểu đồ mũi tên làm nổi bật sự đồng thuận giữa tất cả các bộ dữ liệu ở cấp độ mẫu. f Bản đồ hình ảnh phân cụm của chữ ký đa omics. Các mẫu được đại diện ở các hàng, trong khi các đặc trưng được chọn từ mỗi ba loại omics được đại diện ở các cột. g Biểu đồ circosPlot cho thấy các mối tương quan tích cực (đường màu đỏ) và tiêu cực (đường màu xanh lam) (|r| ≥ 0.5) giữa các đặc trưng được chọn từ mỗi bộ dữ liệu (tên đặc trưng xuất hiện trong mỗi tứ giác).
Một quy trình lựa chọn tiến lên đã xác định rằng các probiotics đối kháng, nối chặt đường ruột, transcriptome của tôm, microbiome đường ruột, và độ ổn định mạng ruột là những yếu tố chính làm giảm độ phong phú của tác nhân gây bệnh (Bảng bổ sung 5). Một mô hình đường dẫn phương pháp phần nhỏ (PLS-PM) cho thấy rằng việc bổ sung probiotics có ảnh hưởng tích cực đến microbiota đường ruột (0.55), độ ổn định mạng (0.56), và nối chặt đường ruột (0.71) (Hình 7a). Thêm vào đó, probiotics tác động tiêu cực trực tiếp (−0.40) lên ba tác nhân Vibrio, cũng như có tác động gián tiếp (−0.51). Trong khi đó, độ ổn định mạng (−0.37, tác động trực tiếp (−0.33) cộng với tác động gián tiếp (−0.04)) và nối chặt đường ruột (−0.41, tác động trực tiếp (−0.31) cộng với tác động gián tiếp (−0.10)) đã tác động tiêu cực đến ba tác nhân Vibrio (Hình 7b). Vì những lý do này, các rào cản sinh thái và động cơ cùng nhau làm giảm sự phát triển của tác nhân gây bệnh. Thật bất ngờ, cấu trúc chức năng tổng thể của đường ruột và transcriptome của tôm đã tác động tiêu cực đến mức độ của các tác nhân mặc dù ảnh hưởng của chúng là không có ý nghĩa (Hình 7a). Đáng chú ý, các tác nhân Vibrio đã mạnh mẽ và tích cực (0.76) gây ra tỷ lệ tử vong ở tôm (Hình 7a). Tóm lại, việc bổ sung probiotics bảo vệ tôm khỏi WFS thông qua việc trực tiếp giảm thiểu tác nhân và gián tiếp tăng cường độ ổn định mạng ruột và nối chặt đường ruột.
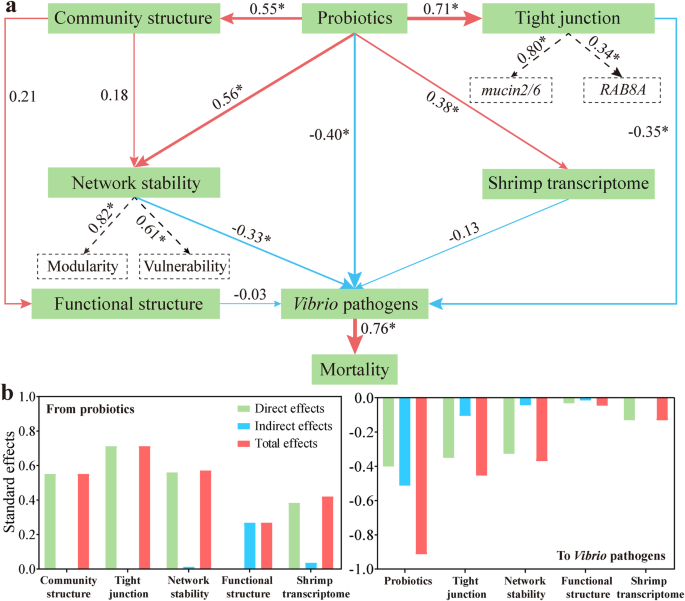
a PLS-PM cho thấy các mối quan hệ chuỗi của các yếu tố khác nhau sau 1000 lần lặp lại. Tải cho độ ổn định mạng đường ruột và nối chặt đường ruột tạo ra các biến tiềm ẩn được thể hiện trong các hình chữ nhật nét đứt. Hệ số đường dẫn tỷ lệ thuận với độ rộng của mũi tên. Màu đỏ và xanh ghi nhận mối quan hệ tích cực và tiêu cực, tương ứng. b Tác động chuẩn hóa của mỗi yếu tố đến tổng độ phong phú của các tác nhân được tính toán từ PLS-PM. Các tác động trực tiếp và gián tiếp được cộng lại để tạo thành tác động tổng hợp.
Nguồn : https://www.nature.com/articles/s41522-024-00509-5


